Oncogenesis
Cancer is a condition in which abnormal cells divide uncontrollably and destroy body tissues. Cell division occurs in almost all tissues to replace worn-out and damaged cells which is why cancer can occur almost everywhere in the body. Oncogenesis is a process in which normal cells are transformed into cancer cells. Oncogenesis is characterized by changes at the cellular, genetic, and epigenetic level that reprograms a cell to undergo uncontrolled cell division leading to malignant mass growths. Oncogenesis is accompanied by the accumulation of multiple mutations. Normally, both proliferation and apoptosis are tightly managed to ensure the integrity of organs and tissues. Mutations and epi-mutations in DNA which disrupt this fine balance between proliferation and apoptosis eventually leads to cancer.
The changes that occur at the cellular, genetic, and epigenetic levels are usually caused by chemical, radiation, or viral induced DNA damage that the cell fails to repair. A mutation in a single gene is not enough to cause cancer but when malignancy develops multiple mutations involving multiple genes accumulate. Tumors arise from clonal growth of cells that have incurred mutations in four classes of genes including proto-oncogenes, tumor suppressor genes, genes that regulate apoptosis, and DNA repair.
Cancer Immunity
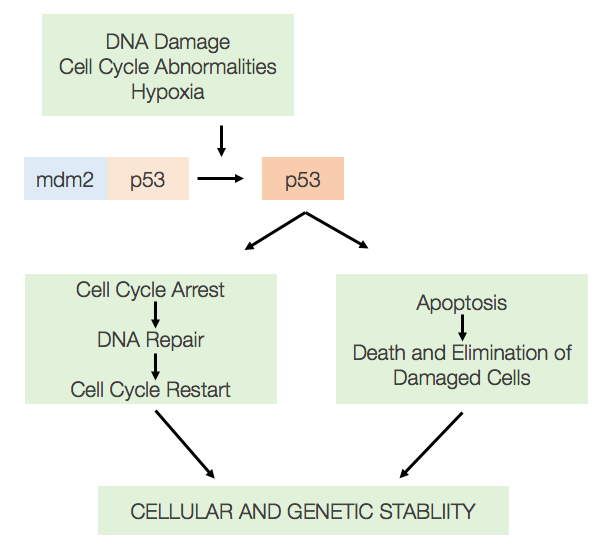
Hundreds of cancer-associated genes have been discovered with p53 being one of the most common mutants. The p53 protein is a gene that codes for a protein that regulates the cell cycle and functions as a tumor suppressor. The p53 protein is directly involved in both growth arrest and apoptosis. DNA damage, cell cycle abnormalities, and hypoxia all can activate the p53 protein. Therefore, an abnormal p53 protein allows oncogenic cells to keep replicating which can lead to the development of cancer.
The immune defense against cancer is a T cell mediated immune response employing the CD8+ Cytotoxic T Lymphocytes (CTL) which identify and lyse the cancer that carry high antigenic mutations by initiating programmed cell death. Cancer cells can survive this immune surveillance through the development of a mechanism which effectively suppress immune responses by activating negative regulatory checkpoints that are associated with self-tolerance or by adopting features that enable them to escape detection. Two such checkpoints that have been most actively researched is the cytotoxic T-lymphocyte protein 4 (CTLA4) and programmed cell death protein 1 (PD-1). CTLA4 is a negative regulator of T cells that raise the threshold for T cell activation. PD-1 is a cell-surface receptor which is expressed by T cells which upon binding to one of two ligands, PD-L1 and PD-L2, suppresses T cell activities. Many types of cells can express PD-L1, including tumor cells and immune cells after exposure to certain interferons. The binding of PD-L1 or PD-L2 to PD-1 generates an inhibitory signal that reduces the activity of T cells. When these checkpoints are suppressed by cancer cells, T cell activation is no longer occurring and the cancer cells can bypass detection by the immune system.
Inside the cancer tissue are special cells which have the characteristics of stem cells and are capable of expressing genes in all cell types. Cancer stem cells can self‐renew, drive tumorigenesis, and metastasis by giving rise to new tumors. There are several transmembrane glycoproteins aberrantly expressed in cancer stem cells which are crucial for cancer to maintain its growth and metastasis although they are highly antigenic. MUC1 is one of such molecules. MUC1 encodes mucin, a highly glycosylated, type I transmembrane glycoprotein normally found in epithelial cells. MUC1 glycoprotein is often found overexpressed with aberrant glycosylation or hypo-glycosylated in a variety of cancers. Its expression is critical for cancer stem cells to maintain ‘stemness’ to promote cancer growth, metastasis, and be resistance to drugs. There are two mechanisms that cancer stem cells employ to avoid being lysed by the T cells. It has been found that epitopes of such aberrant MUC1 protein have been coated with IgG antibodies produced by the cancer cells. Such IgG antibodies do not trigger host immune responses. By masking the critical motifs of the MUC1 molecules, the cancer cells cannot be recognized and lysed by the T cells. The secondary source of IgG antibodies is from the host immune system. Using antibodies to kill cancer is not a productive pathway because it is not strong enough to lyse the cancer stem cells. Rather, their binding to the critical motifs of the aberrant MUC1 causes interference to the T cell action.
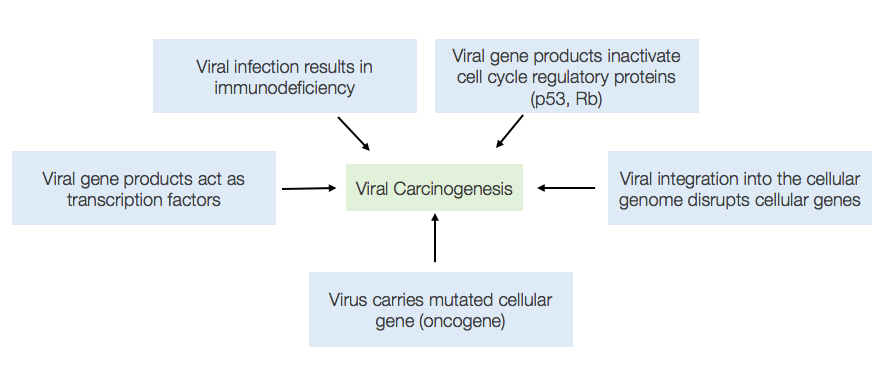
Oncovirus
An oncovirus is a virus that can cause cancer. There are seven viruses that have been identified as an oncovirus including Hepatitis C and B virus, human papillomavirus (HPV), human T-lymphotrophic virus (HTLV-1), Merkel Cell Polyomavirus, Epstein-Barr virus (EBV), Human immunodeficiency virus (HIV), and Human herpesvirus 8 (HHV-8). In most cases, specific viruses only affect specific cells. Viruses use a variety of mechanisms to promote neoplasia (growth of abnormal tissue) through both direct and indirect mechanisms. Most of these viruses either interfere with the p53 protein or are taken up by healthy cells and are copied and replicated which can integrate into the host cell genes. By integrating the viral DNA into host cell chromosomal DNA, they may cause mutations and chromosomal rearrangements that predispose the cell to cancer. The insertion of viral promoter sequences adjacent to certain host genes can lead to ectopic gene expression resulting in neoplasia.
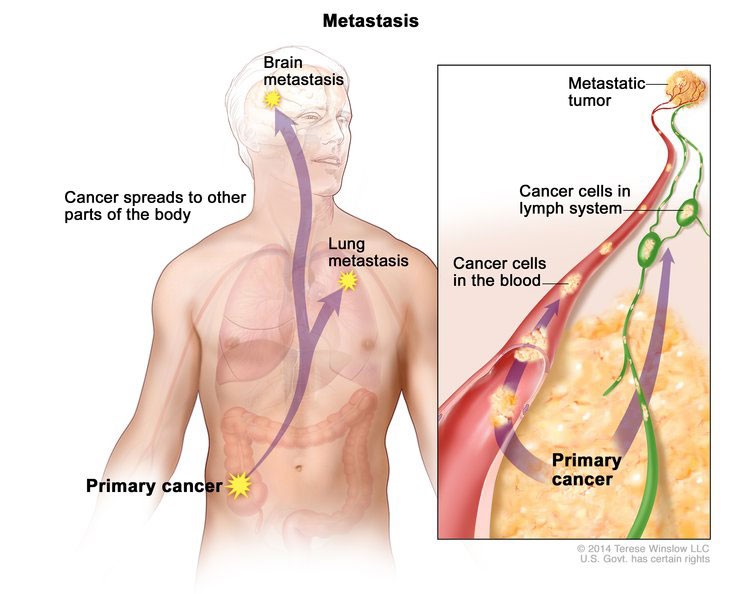
Metastases
The majority of deaths caused by cancer are due to the metastasis of the original tumor cells to sites distant from the primary tumor. Metastasis is the process by which cancer cells migrate throughout the body.
In order for the cancer cells to move through the body, they must first rearrange their cytoskeleton and attach to the other cells and the extracellular matrix via proteins on the outside of their plasma membranes in order to move around neighboring cells. The cells can crawl until they hit a blockage which cannot be bypassed. Often this block is a thick layer of proteins and glycoproteins surrounding the tissues, called the basal lamina or basement membrane. In order to cross this layer, cancer cells secrete a mixture of digestive enzymes that degrade the proteins in the basal lamina and allow them to crawl through.
The proteins secreted by cancer cells contain a group of enzymes called matrix metalloproteases (MMP). These enzymes cut through the proteins that inhibit the movement of the migrating cancer cells. Once the cells have crossed the basal lamina, they can spread through the body in several ways. They can enter the bloodstream or lymphatic system by squeezing between the cells that make up the blood or lymph vessels. Once in the blood stream or in lymphatic circulation, the cells can move through the circulatory system until they find a favorable location to re-enter the tissues. When this occurs, the cancer cells can begin to proliferate to form a new tumor.
If the cancer cells traveled through the lymphatic system, they may end up in the lymph nodes. Usually the lymph nodes near the tumor are affected. These are the nodes that have been doing most of the work to filter out or kill the cancer cells.
Lymph nodes play an important role in cancer staging, which determines the extent of cancer in the body. One of the most commonly used systems for staging cancer is the TNM system, which is based on the extent of the tumor (T), the extent spread to the lymph nodes (N), and the presence of metastasis (M). If there’s no cancer found in the lymph nodes near the cancer, the N is assigned a value of 0. If nearby or distant nodes show cancer, the N is assigned a number (such as 1, 2 or 3), depending on how many nodes are affected, how much cancer is in them, how large they are, and where they are.
Tumors are typically encapsulated by a layer of fibrotic tissue. A benign tumor will grow within the fibrotic connective tissue capsule. However, malignant tumors only remain localized and encapsulated for a short period of time and quickly penetrate the connective tissue to start invade the surrounding tissue. A malignant tumor presents spikes at the surface of the tumor mass.
Tumor Necrosis Factor-α (TNF-α) is a major pro-inflammatory cytokine that are mainly secreted by macrophages. TNF-α can induce apoptosis causing cell death on a certain tumor cell line. However, cancer cells are also capable of producing TNF-α to cause inflammation of its surrounding tissues allowing the spikes to penetrate and invade surrounding tissue and metastasize to other locations.